Hasta taşıma lifti, transfer sürecini daha güvenli ve daha kontrollü hale getirmek için kullanılan önemli bir yardımcı ekipmandır. Ancak bu ekipman tek başına güvenlik sağlamaz. Yanlış kullanım, acelecilik, yetersiz hazırlık veya eğitim eksikliği hem bakım alan kişi hem de bakım veren için ciddi risk yaratabilir. Özellikle yataktan sandalyeye, sandalyeden tuvalet alanına ya da farklı oturma yüzeyleri arasındaki geçişlerde yapılan küçük hatalar büyük sorunlara dönüşebilir. Bu yazıda hasta taşıma lifti kullanırken en sık yapılan hataları, bu hataların neden riskli olduğunu ve daha güvenli kullanım için nelere dikkat edilmesi gerektiğini ele alacağız.
Lifti kullanmadan önce hazırlık yapmamak en temel hatalardan biridir
Hasta taşıma lifti kullanılırken birçok sorun, transfer başlamadan önce yapılan eksik hazırlıklardan kaynaklanır. Oysa güvenli transferin önemli bir kısmı, daha kişi kaldırılmadan önce başlar. Hedef yüzeyin hazır olmaması, tekerlekli sandalye ya da yatak konumunun uygun ayarlanmaması, zeminde engel bulunması veya hareket alanının dar bırakılması transferi zorlaştırır.
Bazı bakım verenler yalnızca liftin çalışıyor olmasına odaklanır. Ancak bu yeterli değildir. Kişinin nereye alınacağı, varış noktasında nasıl yerleştirileceği ve transfer sırasında hangi adımların izleneceği önceden düşünülmelidir. Örneğin sandalye frenlenmemişse, ayaklıklar uygun pozisyona getirilmemişse veya çevrede küçük mobilyalar, halı kıvrımları ya da kablolar varsa, işlem sırasında beklenmedik zorluklar ortaya çıkabilir.
Buradaki temel mantık nettir: Transfer sırasında düşünmek yerine, transferden önce hazırlık yapmak gerekir. Çünkü kişi askıdayken yapılan panik düzeltmeleri hem kullanıcıyı huzursuz eder hem de güvenliği bozar.
Uygun askı ve yerleştirme seçimini hafife almak ciddi risk oluşturur
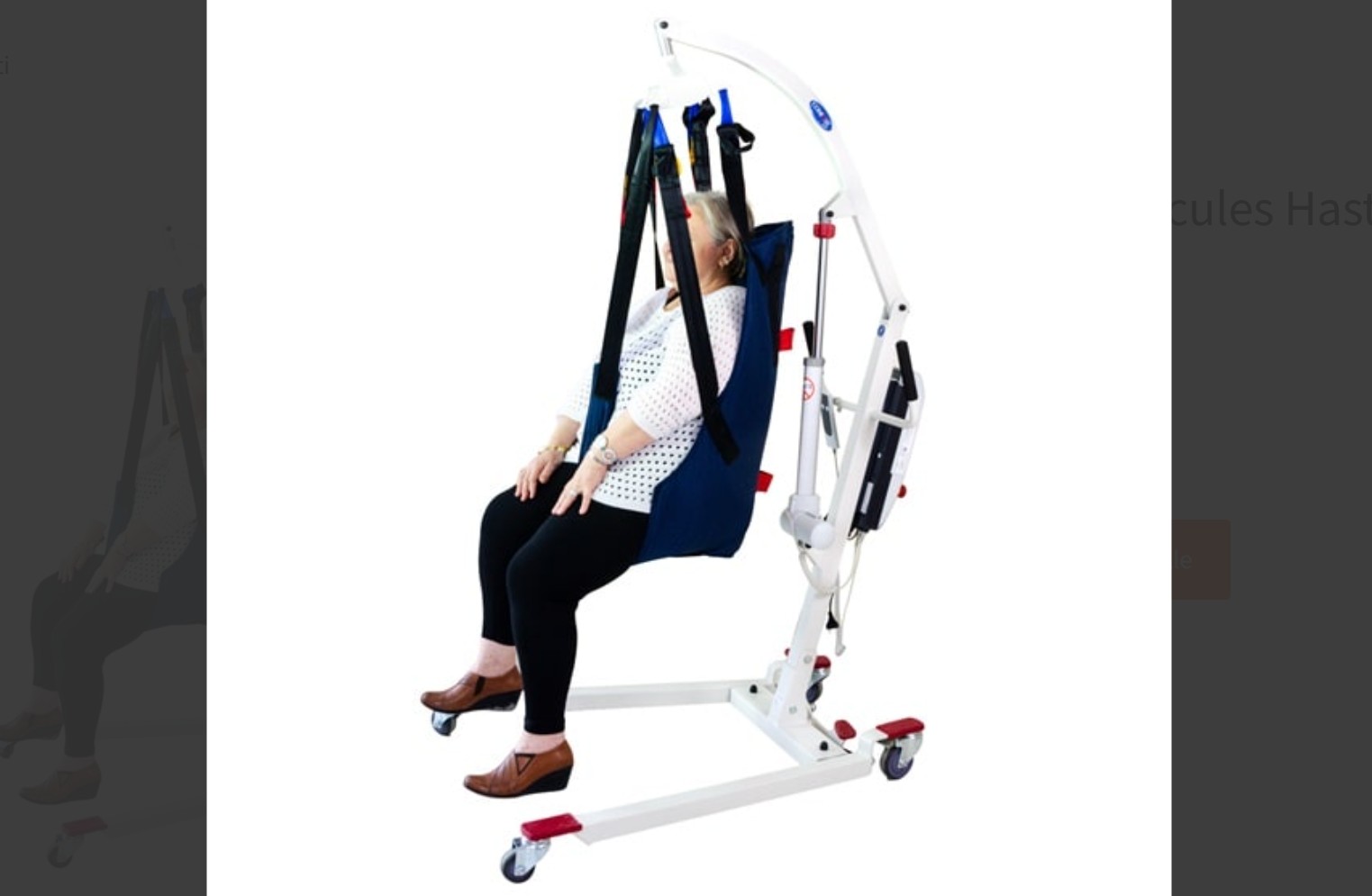
Hasta taşıma liftinde en kritik noktalardan biri askının doğru seçilmesi ve doğru yerleştirilmesidir. Buna rağmen sık yapılan hatalardan biri, askının her kullanıcı için aynı şekilde uygun olduğunu düşünmektir. Oysa kişinin gövde kontrolü, oturma dengesi, destek ihtiyacı ve transfer şekli askı seçiminde belirleyicidir.
Yanlış yerleştirilen askı, kişinin vücut ağırlığını dengeli taşımayabilir. Bu da kaldırma sırasında kayma hissi, dengesizlik ya da rahatsız edici pozisyonlar yaratabilir. Özellikle askı bantlarının yanlış noktaya takılması, askının tam açılmaması veya kişinin vücut altında düzgün yerleştirilmemesi güvenlik açısından büyük hatadır. Kullanıcı askı içinde güvensiz hissettiğinde transferin tamamı daha stresli hale gelir.
Burada “nasıl olsa tutuyor” yaklaşımı yanlıştır. Askının kişiyi sadece taşıması yetmez; dengeli, kontrollü ve rahat şekilde taşıması gerekir. Eğer kullanıcıda baş, gövde ya da kalça desteği ihtiyacı varsa, bu detay daha da önem kazanır. Askı seçimi ve yerleştirme, lift kullanımının teknik ayrıntısı değil, güvenliğin merkezidir.
Transfer sırasında acele etmek ve iletişimi ihmal etmek sık görülen bir sorundur
Hasta taşıma lifti kullanılırken acelecilik, en tehlikeli alışkanlıklardan biridir. Bazı bakım verenler işlemi ne kadar hızlı tamamlarsa o kadar iyi olduğunu düşünür. Oysa hızlı değil, kontrollü transfer güvenlidir. Kişiyi ani şekilde yükseltmek, yön değiştirirken hızlı davranmak ya da hedef yüzeye yaklaşırken sabırsız olmak kullanıcıda hem fiziksel hem psikolojik rahatsızlık yaratabilir.
Transfer sırasında kişiyle iletişim kurmak da önemlidir. Kullanıcı konuşabiliyorsa, hangi adımın yapılacağı önceden söylenmelidir. Bu yaklaşım sadece rahatlatıcı değildir; aynı zamanda kişinin vücut tepkisini daha uyumlu hale getirebilir. Ne zaman kaldırılacağını, hangi yöne dönüleceğini veya nereye indirileceğini bilmeyen biri daha fazla gerilebilir. Bu da transferi zorlaştırır.
Bakım veren açısından da iletişim önemlidir. Eğer transfer iki kişiyle yapılıyorsa, kimin hangi adımı yöneteceği net olmalıdır. Aynı anda farklı hareketler yapmak, koordinasyonu bozabilir. Güvenli kullanım, ekipmanın gücüne değil, sürecin sakin ve planlı yönetilmesine bağlıdır.
Zemini, frenleri ve çevresel koşulları göz ardı etmek güvenliği zayıflatır
Hasta taşıma lifti kullanırken yapılan bir diğer yaygın hata, sadece liftin kendisine odaklanıp çevresel koşulları ihmal etmektir. Oysa zemin yapısı, tekerleklerin hareketi, yatak yüksekliği, sandalye pozisyonu ve çevredeki alan doğrudan transfer güvenliğini etkiler.
Dar alanlarda liftle manevra yapmaya çalışmak, zemindeki küçük engelleri önemsememek veya sandalyeyi uygun pozisyonda sabitlememek transferi riskli hale getirir. Özellikle kaygan yüzeyler, halı kenarları veya eşikler lift hareketini beklenmedik biçimde etkileyebilir. Bu da kaldırma ve indirme sırasında kontrol kaybına yol açabilir.
Ayrıca hedef yüzeyin yüksekliği de önemlidir. Yatağın ya da sandalyenin pozisyonu uygun değilse, son yerleştirme aşaması zorlaşabilir. Bu noktada birçok kişi doğaçlama düzeltme yapmaya çalışır. Asıl kaçınılması gereken budur. Transfer sırasında riskli doğaçlamalara girilmemeli, uygun olmayan koşullarda zorlayıcı hareketler yapılmamalıdır. Bakım verenin güvenli kullanım eğitimi ve temel transfer mantığını bilmesi bu yüzden çok önemlidir.




